|

Seed germination during salt stress |
Sub-functionalization of
Ethylene Receptor Isoforms and Non-Canonical Signaling
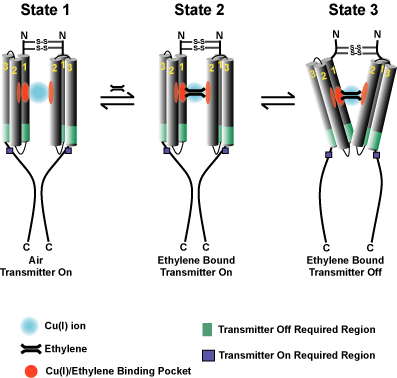
A third area of current research is to determine the basis for
ethylene receptor sub-functionalization and non-canonical signaling pathways (reviewed in
Shakeel et al., 2013;
Binder, 2020). We have uncovered several
instances where the receptors have become sub-functionalized. For
instance, ETR1, ETR2, and EIN4 are required for normal growth recovery
after removal of ethylene, while ERS1 and ERS2 are not. This function
requires ETR1 histidine kinase activity which seems to signal to the cytokinin pathway (Binder
et al., 2004; Binder
et al. 2018). ETR1 plays the predominant role in mediating the
inhibitory effects of silver ions on ethylene responses (McDaniel
and Binder, 2012), in
mediating many responses of roots to ethylene
(Harkey et al.,
2018) and in susceptibility to
the cyst nematode
Heterodera schactii (Piya et al.,
2019). Interestingly, our research has also revealed instances of
contrasting roles for certain receptor isoforms. The first instance that we
found was that ETR1 is necessary and sufficient for ethylene-stimulated nutational bending (also called circumnutations) of Arabidopsis
hypocotyls; by contrast loss of the other four receptor isoforms leads to constitutive nutations (Binder
et al., 2006;
Kim et al., 2011).
Another instance of contrasting roles is that ETR1 (and to a lesser extent EIN4) inhibits and ETR2
stimulates seed germination during salt stress or in the presence of ABA and in darkness (Wilson
et al., 2014a;
Wilson
et al., 2014b; Bakshi et al., 2018). The receiver domain of ETR1 has an important
role during germination under salt stress. We've recently defined regions of the
receiver domain important for specific traits (Bakshi
et al., 2015).
Thus, we have two instances where receptor isoforms have opposite roles
in a trait. This is not explained by current models of ethylene
signaling. We are now studying the mechanistic basis
for these various roles. |



 and
and